The cell can be likened to a specialized computer that executes the DNA program and expresses the information contained in it. Cats, dogs, horses, and Archaeopteryxes don't really evolve, of course, but live their spans and die still being genetically pretty much the same as they were when born. What evolves, according to the theory, is the package of genetic information that gets passed down from generation to generation, accumulating and preserving beneficial innovations as it goes. The species that exists at a given time is a snapshot of the genome expressing itself as it stands at the point it has reached in accumulating information down the line of descent from the earliest ancestor. Although the process may be rapid at times and slow at others, every mutation that contributes to the process adds something on average. This is another way of saying that to count as a meaningful evolutionary step, a mutation must add some information to the genome. If it doesn't, it contributes nothing to the building up of information that the evolution of life is said to be.
No mutation that added information to a genome has ever been observed to occur, either naturally or in the laboratory. This is the crucial requirement that disqualifies all the examples that have been presented in scientific papers, reproduced in textbooks, and hyped in the popular media as "evolution in action." We already saw that the case of the peppered moth involves no genetic innovation; what it demonstrates is an already built-in adaptation capacity, not evolution. This isn't to say that mutations never confer survival benefits in some circumstances. Such occurrences are rare, but they do happen. However, every one that has been studied turns out to be the result of information being lost from a genome, not gained by it. So what's going on in such situations is just part of the normal process of existing organisms shuffling and jostling in their own peculiar ways for a better place in the sun, but not turning into something new.
A frequently cited example is that of bacteria gaining resistance to streptomycin and some other mycin drugs, which they are indeed able to do by a single-point mutation. The drug molecule works by attaching to a matching site on a ribosome (protein-maker) of the bacterium, rather like a key fitting into a lock, and interfering with its operation. The ribosome strings the wrong amino acids together, producing proteins that don't work, as a result of which the bacterium is unable to grow, divide, or propagate, and is wiped out. Mammalian ribosomes don't have similar matching sites for the drug to attach to, so only the bacteria are affected, making such drugs useful as antibiotics. However, several mutations of the bacterial genome are possible that render the drug's action ineffective. In a population where one of them occurs, it will be selected naturally to yield a resistant strain which in the presence of the antibiotic indeed has a survival benefit.
But the "benefit" thus acquired turns out to be a bit like gaining immunity to tooth decay by losing your teeth. Every one of the resistance-conferring mutations does so by altering one part or another of the ribosome "lock" in such a way that the drug's molecular "key" will no longer match. This is another way of saying that the specific set of lock parts that enables the key fit is replaced by one of several randomly determined alternative sets that it won't fit. The significant point is that a single, unique state is necessary to bring about the first condition, "key fits," whereas any one of a number of states is sufficient to produce the second condition, "key doesn't fit." Thinking of it as a combination lock, only one combination of all digits will satisfy the first condition, but altering any digit (or more) meets the second. This makes a number less specificsuch as by changing 17365 to 173X5, where X can be any digit. Loss of specificity means a loss of information. The same applies to pests becoming resistant to insecticides such as DDT. Although a survival benefit may be acquired in certain circumstances, the mutant strains invariably show impairment in more general areas, such as by slowed metabolism or sluggish behavior. Hence, they turn out to be not "super species" at all, as the media love to sensationalize, but genetic degenerates which if the artificial conditions were taken away would rapidly be replaced by the more all-round-rugged wild types.
Losing the genes that control the growth of teeth might produce a strain of survivors in a situation where all the food that required chewing was poisoned and only soup was safe. But it couldn't count as meaningful in any evolutionary sense. If evolution means the gradual accumulation of information, it can't work through mutations that lose it. A business can't accumulate a profit by losing money a bit at a time.
Neither can it do so through transactions that break even. Some bacteria can become resistant through infection by a virus carrying a gene for resistance that the virus picked up from a naturally resistant variety. Some insects seem to get their uncannily effective camouflage by somehow having acquired the same color-patterning genes as are possessed by the plants they settle on. 29 Similar results can also be achieved artificially by genetic engineering procedures for transferring pieces of DNA from one organism to another. Although it is true that information is added to the recipient genomes in such cases, there is no gain for life as a whole in the sense of a new genetic program being written. The program to direct the process in question was already in existence, imported from somewhere else. Counting it as contributing to the evolution of life would be like expecting an economy to grow by having everyone take in everyone else's laundry. For an economy to grow, wealth must be created somewhere. And as we've seen, considerations of the probabilities involved, limitations of the proposed mechanisms, and all the evidence available, say that theories basing large-scale evolution on chance don't work.
Cases of adaptations occurring not through selection of random changes but being directed by cues in the environment have been reported for over a century. 30 But since any suggestion of nonrandom variation goes against the prevailing beliefs of mainstream biology, they have largely been ignored. Take, for example, the backup feeding system that the laboratory staple bacterium E. coli is able to conjure up on demand. 31
The normal form of E.coli lives on the milk sugar lactose and possesses a set of digestive enzymes tailored to metabolize it. A defective strain can be produced that lacks the crucial first enzyme of the set, and hence cannot utilize lactose. However, it can be raised in an alternative nutrient. An interesting thing now happens when lactose is introduced into the alternative nutrient. Two independent mutations to the bacterium's genome are possible which together enable the missing first step to be performed in metabolizing lactose. Neither mutation is any use by itself, and the chances of both happening together is calculated to be vanishingly small at 10-18. For the population size in a typical experiment, this translates into the average waiting time for both mutations to happen together by chance being around a hundred thousand years. In fact, dozens of instances are found after just a few days. But only when lactose is present in the nutrient solution. In other words, what's clearly indicated in experiments of this kindand many have been described in the literature 32 is that the environment itself triggers precisely the mutations that the organism needs in order to exploit what's available.
The forms of adult animal bone-muscle systems are influenced to a large degree by the forces that act on them while they are growing. Jaws and teeth have to bear the forces exerted when the animal chews its food, and these forces will depend in strength and direction on the kind of food the animal eats. The adult form of jaws and teeth that develops in many rodents, for example, can vary over wide ranges with changes in diet, brought about possibly by environmental factors or through a new habit spreading culturally through a population. If the new conditions or behavior become established, the result can be a permanent change in the expressed phenotype of the animal.
In 1967, a hundred or so finches of the same species were brought from Laysan, an island in the Pacific about a thousand miles northwest of Hawaii, forming part of a U.S. government bird reservation, to a small atoll called Southeast Island, somewhat southeast of Midway, which belongs to a group of four small islands all within about ten miles of each other. Twenty years later, the birds had dispersed across all the islands and were found to have given rise to populations having distinct differences, particularly with regard to the shapes and sizes of their beaks. 33 Clearly this wasn't the result of randomly occurring mutations being naturally selected over many generations. The capacity to switch from one form to another was already present in the genetic program, and the program was switched to the appropriate mode by environmental signals. The ironic aspect of this example, of course, is that observations of precisely this type of variety in beak forms among finches of the Galapagos Islands led Darwin to the notion that he was witnessing the beginnings of new species.
By the above, if a population of rodents, say, or maybe horses, were to shift their diet abruptly, the phenotype would change abruptly even though the genotype does not. The fossil record would show abrupt changes in tooth and bone structure, even though there had been no mutation and no selection. Yet the evolution read into the fossil record is inferred largely from bones and teeth. In his reconstruction of the story of horse evolution, Simpson tells that when the great forests gave way to grassy plains, Mesohippus evolved into Merychippus, developing high-crowned teeth through random mutation and selection, for "It is not likely to be a coincidence that at the same time grass became common, as judged by fossil grass seeds in the rocks." 34
It may indeed have been no coincidence. But neither does it have to be a result of the mechanism that Simpson assumes. If these kinds of changes in fossils were cued by altered environments acting on the developing organisms, then what has been identified as clear examples of evolution could have come about without genetic modification being involved, and with random mutation and selection playing no role at all.
Should this really be so strange? After all, at various levels above the genetic, from temperature regulation and damage repair to fighting or fleeing, organisms exhibit an array of mechanisms for sensing their environment and adjusting their response to it. The suggestion here is that the principle of sensing and control extends down also to the genetic level, where genes can be turned on and off to activate already-existing program modules, enabling an organism to live efficiently through short-term changes in its environment. Nothing in the genome changes. The program is set up for the right adaptive changes in the phenotype to occur when they are needed.
The problem for Darwinism, and maybe the reason why suggestions of directed evolution are so fiercely resisted, is that if there was trouble enough explaining the complexity of genetic programs before, this makes it immeasurably worse. For now we're implying a genome that consists not only of all the directions for constructing and operating the self-assembling horse, but also all the variations that can be called up according to circumstances, along with all the reference information to interpret the environmental cues and alter the production specification accordingly. Fred Hoyle once observed that the chances of life having arisen spontaneously on Earth were about on a par with those of a whirlwind blowing through a junkyard containing all the pieces of a 747 lying scattered in disarray, and producing an assembled aircraft ready to fly. What we're talking about now is a junkyard containing parts for the complete range of Boeing civil airliners, and a whirlwind selecting and building just the model that's best suited to the current situation of cost-performance economics and projected travel demands.
So finally we arrive at the reason why the subject is not just a scientific issue but has become such a battle of political, moral, and philosophic passions. At the root of it all, only two possibilities exist: Either there is some kind of intelligence at work behind what's going on, or there is not. This has nothing to do with the world's being six thousand years old or six billion. A comparatively young worldin the sense of the surface we observe todayis compatible with unguided Catastrophist theories of planetary history, while many who are of a religious persuasion accept orthodox evolution as God's way of working. What's at the heart of it is naturalism and materialism versus belief in a creative intelligence of some kind. Either these programs which defy human comprehension in their effectiveness and complexity wrote themselves accidentally out of mindless matter acting randomly; or something wrote them for a reason. There is no third alternative.
At the time Darwin formulated his original theory, nothing was known of the mechanism of heredity or the internal structures of the organic cell. The cell was known to possess a dark nucleus, but the inner workings were pretty much a "black box," imagined to be a simple unit of living matter, and with most of the interesting things taking place at higher levels of organization. With the further development of sciences leading to the molecular biology that we see today, this picture has been dramatically shattered and the cell revealed as the stupendous automated factory of molecular machines that we glimpsed in Michael Denton's description earlier. The complexity that has been revealed in the last twenty years or so of molecular biochemistry is of an order that dwarfs anything even remotely imagined before then.
These findings prompted Michael Behe, professor of biochemistry at Lehigh University in Pennsylvania, to write what has become an immensely popular and controversial book, Darwin's Black Box, 35 in which he describes systems ranging from the rotary bearings of the cilia that propel mobile cells, to vision, the energy metabolism, and the immune system, which he argues cannot have come into existence by any process of evolution from something simpler. His basis for this assertion is the property they all share, of exhibiting what he terms "irreducible complexity." The defining feature is that every one of the components forming such a system is essential for its operation. Take any of them away, and the system is not merely degraded in some way but totally incapable of functioning in any way at all. Hence, Behe maintains, such systems cannot have arisen from anything simpler, because nothing simplerwhatever was supposed to have existed before the final component was addedcould have done anything; and if it didn't do anything, it couldn't have been selected for any kind of improvement. You either have to have the whole thingwhich no variation of evolution or any other natural process could bring into existence in one stepor nothing.
The example he offers to illustrate the principle is the common mousetrap. It consists of five components: a catch plate on which the bait is mounted; a holding bar that sets and restrains the hammer; a spring to provide the hammer with lethal force; and a platform for mounting them all on and keeping them in place. Every piece is essential. Without any one, nothing can work. Hence, it has to be built as a complete, functioning unit. It couldn't assume its final form by the addition of any component to a simpler model that was less efficient.
An example of reduced complexity would be a large house built up by additions and extensions from an initial one-room shack. The improvements could be removed in reverse order without loss of the essential function it provides, though the rendering of that function would be reduced in quality and degree.
Here, from Behe's book, are the opening lines of a section that sketches the process of vision at the biochemical level. Nobody has been able to offer even a speculation as to how the system could function at all if even one of its molecular cogs were removed.
When light first strikes the retina a photon interacts with a molecule called 11-cis-retinal, which rearranges within picoseconds [a picosecond is about the time light takes to cross the width of a human hair] to trans-retinal. The change in the shape of the retinal molecule forces a change in the shape of the protein rhodopsin, to which the retinal is tightly bound. The protein's metamorphosis alters its behavior. Now called metarhodopsin II, the protein sticks to another protein, called transducin. Before bumping into metarhodopsin II, transducin had tightly bound a small molecule called GDP. But when transducin interacts with metarhodopsin II, the GDP falls off, and a molecule called GTP binds to transducin.
Concluding, after three long, intervening paragraphs of similar intricacy:
Trans-retinal eventually falls off rhodopsin and must be reconverted to 11-cis-retinal and again bound by rhodopsin to get back to the starting point for another visual cycle. To accomplish this, trans-retinal is first chemically modified by an enzyme called trans-retinola form containing two more hydrogen atoms. A second enzyme then converts the molecule to 11-cis-retinol. Finally, a third enzyme removes the previously added hydrogen atoms to form 11-cis-retinal, a cycle is complete. 36
The retinal site is now ready to receive its next photon.
Behe gives similarly comprehensive accounts of such mechanisms as blood clotting and the intracellular transport system, where the functions of all the components and their interaction with the whole are known in detail, and contends that only purposeful ordering can explain them. In comparison, vague, less precisely definable factors such as anatomical similarities, growth of embryos, bird lineages, or the forms of horses become obsolete and irrelevant, more suited to discussion in Victorian drawing rooms.
The response from the evolutionists to these kinds of revelations has been almost complete silence. In a survey of thirty textbooks of biochemistry that Behe conducted, out of a total of 145,000 index entries, just 138 referred to evolution. Thirteen of the textbooks made no mention of the subject at all. As Behe notes, "No one at Harvard University, no one at the National Institutes of Health, no member of the National Academy of Sciences, no Nobel prize winnerno one at all can give a detailed account of how the cilium, or vision, or blood clotting, or any other complex biochemical process might have developed in a Darwinian fashion." 37
Behe unhesitatingly sees design as the straightforward conclusion that follows from the evidence itselfnot from sacred books or sectarian beliefs. He likens those who refuse to see it to detectives crawling around a body lying crushed flat and examining the floor with magnifying glasses for clues, while all the time ignoring the elephant standing next to the bodybecause they have been told to "get their man." In the same way, Behe contends, mainstream science remains doggedly blind to the obvious because it has fixated on finding only naturalistic answers. The simplest and most obvious reason why living systems should show over and over again all the signs of having been designedis that they were.
Others whom we have mentioned, such as Denton, Hoyle, Spetner, express similar sentimentsnot through any prior convictions but purely from considerations of the scientific evidence. Interest in intelligent design has been spreading in recent years to include not just scientists but also mathematicians, information theoreticians, philosophers, and others dissatisfied with the Darwinian theory or opposed to the materialism that it implies. Not surprisingly, it attracts those with religious interpretations too, including fundamentalists who insist on a literal acceptance of Genesis. But it would be a mistake to characterize the whole movement by one constituent group with extreme views in a direction that isn't really relevant, as many of its opponents try to doin the same way that it would be to belittle the notion of extraterrestrial intelligence because UFO abduction believers happen to subscribe to it. As Phillip Johnson says, "ID is a big tent" that accommodates many diverse acts. All that's asserted is that the evidence indicates a creative intelligence of some kind. In itself, the evidence says nothing about the nature of such an intelligence nor what its purpose, competence, state of mind, or inclination to achieve what we think it should, might be.
The argument is sometimes put forward that examples of the apparent lack of perfection in some aspects of biological function and adaptation mean that they couldn't be the work of a supreme, all-wise, all-knowing creator. This has always struck me as curious grounds for scientists to argue on, since notions of all-perfect creators were inventions of opponents more interested in devising means for achieving social control and obedience to ruling authorities than interpreting scientific evidence. Wrathful gods who pass judgments on human actions and mete out rewards or retribution make ideal moral traffic policemen, and it seems to be only a matter of time (I put it at around 200-300 years) before religions founded perhaps on genuine insights for all I know are taken over by opportunists and sell out to, or are coopted by, the political power structure. In short, arguments are made for the reality of some kind of creative intelligence; human social institutions find that fostering belief in a supreme moral judge is to their advantage. Nothing says that the two have to be one and the same. If the former is real, there's no reason why it needs to possess attributes of perfection and infallibility that are claimed for the latter. Computers and jet planes are products of intelligence, but nobody imagines them to be perfect.
Those who are persuaded by religious interpretations insist on the need for a perfect God to hand down the absolute moral standards which they see as the purpose in creating the worldand then go into all kinds of intellectual convolutions trying to explain why the world clearly isn't perfect. I simply think that if such an intelligence exists it would do things for its reasons not ours, and I don't pretend to know what they might bealthough I could offer some possibilities. An analogy that I sometimes use is to imagine the characters in a role-playing game getting complex enough to become aware that they were in an environment they hadn't created, and which they figure couldn't have created itself. Their attempts to explain the reason for it all could only be framed in terms of the world that they know, that involves things like finding treasures and killing monsters. They could have no concept of a software writer creating the game to meet a specification and hold down a job in a company that has a budget to meet, and so on.
I sometimes hear the remark that living things don't look like the products of design. True enough, they don't look very much like the things we're accustomed to producing. But it seems to me that anyone capable of getting self-assembling protein systems to do the work would find better things to do than spend their existence bolting things together in factories. Considering the chaotically multiplying possibilities confronting the development of modules of genetic code turned loose across a range of wildly varying environments to make what they can of themselves, what astounds me is that they manage as well as they do.
These are all valid enough questions to ask, and we could spend the rest of the book speculating about them. But they belong in such realms of inquiry as theology and philosophy, not science.
How confident can we be that design is in fact the necessary explanation, as opposed to some perhaps unknown natural processpurely from the evidence? In other words, how do you detect design? When it comes to nonliving objects or arrangements of things, we distinguish without hesitation between the results of design and of natural processes: a hexagonal, threaded nut found among pebbles on a beach; the Mount Rushmore monument as opposed to a naturally weathered and eroded rock formation; a sand castle on a beach, distinguished from mounds heaped by the tide. Exactly what is it that we are able to latch on to? If we can identify what we do, could we apply it to judging biological systems? William Dembski, who holds doctorates in mathematics and philosophy from the Universities of Chicago and Illinois, has tackled the task of setting out formally the criterion by which design is detected. 38 His analysis boils down to meeting three basic conditions.
The first is what Dembski terms "contingency": that the system being considered must be compatible with the physics of the situation but not required by it. This excludes results that follow automatically and couldn't be any other way. Socrates, for example, believed that the cycles of light and darkness, or the progressions of the seasons pointed toward design. But what else could follow day except night? What could come after cold but warming, or after drought other than rain?
Second is the condition that most people would agree, that of "complexity," which is another way of describing a situation that has a low probability of occurring. Of all the states that the components of a watch might assume from being thrown in a pile or joined together haphazardly, if I see them put together in precisely the configuration necessary for the watch to work, I have no doubt that someone deliberately assembled them that way.
But complexity in itself isn't sufficient. This is the point that people whom I sometimes hear fromand others writing in books, who should know bettermiss when they argue that the information content of a genome is nothing remarkable, since there's just as much information in a pile of sand. It's true that spelling out the position and orientation of every sand grain to construct a given pile of sand would require a phenomenal amount of information. In fact it would be a maximum for the number of components involved, for there's no way of expressing a set of random numbers in any shorter form such as a formula or the way a computer program of a few lines of code could be set up to generate, say, all the even numbers up to ten billion. But the only thing the numbers would be good for is to reconstruct that specific pile of sand. But the specificity means nothing, since for the purposes served by a pile of sand on the ground, one pile is as good as another and so you might as well save all the bother and use a shovel. But the same can't be said of the sequences of DNA base pairs in a genome.
Suppose someone comes across a line of Scrabble tiles reading METHINKS IT IS LIKE A WEASEL, with spaces where indicated. Asked to bet money, nobody would wager that it was the result of the cat knocking them out of the box or wind gusting through the open window. Yet it's not the improbability of the arrangement that forces this conclusion. The sequence is precisely no more or no less probable than any other of twenty-eight letters and spaces. So what is it? The typical answer, after some chin stroking and a frown, is that it "means something." But what does that mean? This is what Dembski was possibly the first to recognize and spell out formally. What we apprehend is that the arrangement, while not only highly improbable, specifies a pattern that is intelligible by a convention separate from the mere physical description. Knowledge of this conventionDembski calls this "side information"enables the arrangement to be constructed independently of merely following physical directions. In this case the independent information is knowledge of the English language, Shakespeare, and awareness of a line spoken by Hamlet. Dembski's term for this third condition is "specificity," which leads to "specified complexity" as the defining feature of an intelligently contrived arrangement.
Specifying a pattern recognizable in English enables the message to be encoded independently of Scrabble tiles, for example into highly improbable configurations of ink on paper, electron impacts on a screen, magnetic dots on a VHS sound track, or modulations in a radio signal. Michael Behe's irreducible complexity is a special case of specified complexity, where the highly improbable organizations of the systems he describes specify independent patterns in the form of unique, intricate biological processes that the components involved, like the parts of a watch, could not perform if organized in any other way.
A process that Richard Dawkins terms "cumulative complexity" is frequently put forward as showing that Darwinian processes are perfectly capable of producing such results. An example is illustrated in the form of a contrived analogy given by the philosopher Elliott Sober that uses the same phrase above from Hamlet. 39 The letters are written on the edges of randomly spun disks, one occupying each position of the target sentence like the wheels of a slot machine. When a wheel happens to come up with its correct letter it is frozen thereafter until the sentence is complete. Ergo, it is claimed, pure randomness and selection can achieve the required result surprisingly rapidly. The idea apparently comes from Richard Dawkins and seems to have captured the imagination of philosophers such as Michael Ruse and Daniel Dennett, who also promote it vigorously.
But their enthusiasm is hard to understand, for the model shows the opposite of what it purports to. Who is deciding which disks to freeze, and why? What the analogy demonstrates is an intelligence directing the assembly of a complex system toward a preordained target already constructed independently of the mechanics by other meansin this case the creativity of Shakespeare. Yet the whole aim of Darwinism was to produce a nonteleological explanation of life, i.e., one in which purpose played no role. Hence, either these advocates don't understand their own theory, or they fail to register that they've disproved their assumptions.
Given that little if anything in life is perfect, how confident could we be in a test using these principles to detect the signature of intelligence in nature? As with a medical test it can err in two ways: by giving a "false positive," indicating design when there isn't any, or a "false negative," by failing to detect design when it was actually present.
We live with false negatives all the time. When the information available is simply insufficient to decidea rock balanced precariously on another; a couple of Scrabble tiles that happen to spell IT or SOour tendency is to favor chance, since the improbabilities are not so high as to rule it out, but we're sometimes wrong. Such instances are specific, yes, but not complex enough to prove design. Intelligence can also mimic natural processes, causing us to let pass as meaningless something encrypted in an unrecognized code or to accept as an accident what had been set up to appear as such when in fact it was arson or a murder. Although we have entire professions devoted to detecting such false negatives, such as police detectives, forensic scientists, and insurance claim investigators, we can get by with imperfection.
False positives are another thing entirely. A test that can discern design where there is none is like reading information into entrails, tea leaves, or flights of birds that isn't there, which makes the test totally useless. Hence, a useful test needs to be heavily biased toward making false negatives, rejecting everything where there's the slightest doubt and claiming a positive only when the evidence is overwhelming. Thinking of it as a net, we'd rather it let any number of false negatives slip through. But if it catches something, we want to be sure that it's a real positive. How sure can we be?
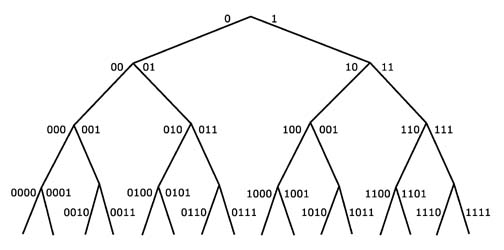
What the criterion of specified complexity is saying is that once the improbabilities of a situation become too vast (2728 possible combinations of the Scrabble example above), and the specification too tight (one line from Hamlet), chance is eliminated as a plausible cause, and design is indicated. Just where is the cutoff where chance becomes unacceptable? The French mathematician Emile Borel proposed 10-50 as a universal probability bound below which chance could be precludedin other words a specified event as improbable as this could not be attributed to chance. 40 This is equivalent to saying it can be expressed in 166 bits of information. How so? Well, Imagine a binary decision tree, where the option at each branch point is to go left or right. The first choice can be designated by "0" or "1," which is another way of saying it encodes one bit of information. Since each branch leads to a similar decision point, the number of branches at the next level will be four, encoded by two bits: 00, 01, 10, and 11. By the time the tree gets to 166 levels, it will have sprouted 1050 branches. The information to specify the path from the starting point to any one of the terminal points increases by one bit for each decision and hence can be expressed as a binary number of 166 bits.
The criterion that Dembski develops applies a bound of 10-150. That's 100 zeros more stringent than the limit beyond which Borel said chance can be discounted. This translates into 500 bits of information. 41 According to Dembski's criterion, specified information of greater than 500 bits cannot be considered as having come about via chance processes. The bacterial cilium that Behe presents as one of his cases of irreducible complexity is a whiplike rotary paddle used for propulsion, driven by an intricate molecular machine that includes an acid-powered engine, stator housing, O-rings, bushings, and a drive shaft, and is built from over 40 interacting proteins, every one of them essential. Its complex specified information is well above 500 bits. So are those of all the other cases Behe gives. And we've already come across improbabilities that are way beyond this bound, such as Fred Hoyle's figure for the likelihood of the full complement of human proteins arising through chance, or Lee Spetner's for speciation and convergence. Many other examples could be cited.
But those who commit a priori to a philosophy that says the universe consists of nothing but matter and motion must accept evolution. The worldview that they have stated as fact leaves no alternative. Things like fossils, genetics, probabilities, and complexity have no real bearing apart from a need for being somehow interpreted to fit, because the issue has already been decided independently of any evidence.
So, to repeat what we said above, either mindless, inanimate matter has the capacity to organize itself purposelessly into the things we've been talking about, or some kind of intelligence caused it to be organized. Now let's go back to the question posed right at the beginning. Based on what we see today, which belief system constrains permissible answers only to those permitted by a prespecified dogma, and which simply follows the evidence, without prejudice, to wherever it seems to be leading? Which, in other words, is the religion, and which is the science?
Some defenders of the Darwinist view evade the issue by defining science as the study of naturalistic, materialistic phenomena and the search for answers to all things only in those terms. But what if the simple reality is that some questions don't have answers in those terms? One response is that science could only be enriched by abandoning that restrictive philosophy and opening its horizons in the way the spirit of free inquiry was supposed to. The alternative could be unfortunate. For in taking such a position, science could end up excluding itself from what could well be some of the most important questions confronting us.
1 Himmelfarb, 1962
2 Dennett, 1995, p. 46
3 New York Times, April 9, 1989, Sec 7, p. 34
4 Dennett, 1995, pp. 515-516
5 Hogan, 1977
6 Hogan, 1988
7 Darwin, 1859, p.184
8 The Origin of Species, 1872, 6th edition, John Murry,
London, p. 4689 The Origin of Species, 1872, 6th edition, John Murray,
London, p. 30910 Sunderland, 1998
11 Denton, 1985, p. 190
12 Johnson, Phillip, 1991, p.51
13 Wells, 2000, Chapter 6
14 Sunderland, 1998, p. 86
15 Johnson, 1991, p. 79
16 See, for example, Sunderland, 1998, p. 94
17 Stanley, 1979, p. 39
18 Dawkins, 1986, p. 1
19 Macbeth, 1971, p. 5
20 According to Simpson, "anything tending to produce
systematic, heritable change in populations between one
generation and the next." Quoted in Macbeth, 1971, p. 4821 Macbeth, 1971, p. 48
22 See Wells, 2000, Chapter 7 for more details and a
discussion on the implications of needing to falsify
textbooks when we're assured that the evidence for
evolution is "overwhelming." A full account of the story
is available online at the Nature Institute,
http://www.netfuture.org/ni/misc/pub/moth.html23 Spetner, 1997, p. 63
24 Judson, 1979, p. 217
25 Denton, 1985, pp. 328-29
26 Spetner, 1997, p. 92
27 Hoyle, 1983, pp. 12-17
28 Sunderland, 1996, p. 152
29 Hoyle, 1983, Chapter 5
30 For examples of reviews see Ho and Saunders, 1979; Cook, 1977; Rosen and Buth, 1980
31 Original research reported in Hall, 1982
32 See Spetner, 1997, Chapter 7 for more examples
33 Spetner, 1997, p. 204
34 Simpson, 1951, p. 173
35 Behe, 1996
36 Behe, 1996, p. 20
37 Behe, 1996. p. 187
38 Dembski 1998, 1999, 2002
39 Sober, 1993
40 Borel, 1962, p. 28
41 Dembski, 1998, Section 6.5